
To understand the physical origin of Delayed Luminescence (DL), we studied the DL emission of condensed systems. We observed a correlation between DL seams and the more o less organised structure of the condensed systems and that it arises form the presence of collective excitations inside the structure such as excitons. We measured DL emission of various types of crystals, in particular molecular crystals, in which there should be the presence of localised excitos or Frenkel excitons (Phenyl Salicytate in solid and liquid states), and crystals of semiconductors, in which there has been hypothesised the existence of delocalised excitons or Wannier-Mott excitons (polycristalline Cadmium Sulphide, Silicon monocrystal). The experiments have showed as the DL is determined essentially by the structure of the systems and disappears if the structure is destroyed. Figure 1 shows the total counts of re-emitted photons by polycristalline Cadmium Sulphide (CdS) as function of grain average dimensions.
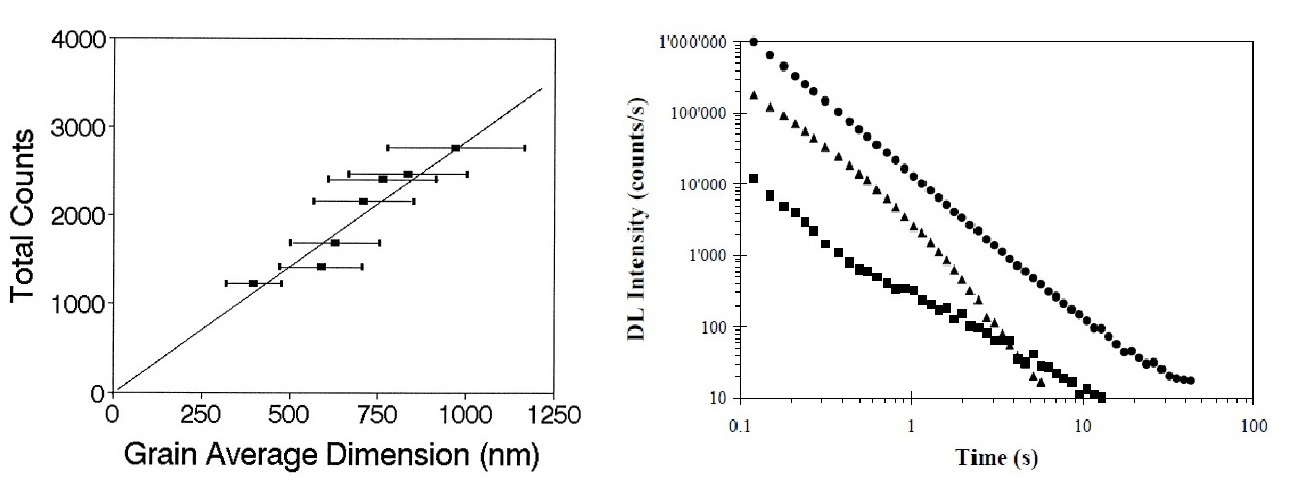
Figure 1. - (on the left) Total counts, i.e total number of emitted photons, as a function of the avegare dimension of grains for CdS samples (J Photochem Photobiol B 56 (2000)). (on the right) DL temporal decays of Cadmium Sulfide polycrystalline samples of different grain sizes: (square) less than 1 mm, (triangle) of the order of 10 mm, (circle) larger than 100 mm. The intensity decreases rapidly (in the first approssimation is linear, with r2=0.97) with the reduction of the dimension of the grains. However the signal is even well measurable fro grains having dimension of the order of hundreed of nm, comparable to the dimension of the ordered structures existing inside the cells.
The numerous experimental data accumulated on DL emission of biological systems and of condensed matter showed the strong correlation between DL properties and the all state of the system under study. For example, the measures carried out on the sulphide Cadmium (a polycrystalline material), and on the seaweed Acetabularia acetabulum (a unicellular organism of large dimensions and characterized by almost one-dimensional ordered structures constituting the cytoskeleton) had shown that the disintegration of the structures (in the case of cadmium) or to their damage (in the case of the alga) there is a decrease in the emission of DL, no drastic change in fluorescence emission.
 Obviously biosystems posses specific delayed photoluminescence whose properties differ significantly from those of similar devitalised organisms, and both are different from non-living systems and depend on the physical, chemical, physiological and other conditions and pre-history of the samples. All our experimental results show that the emission mechanism is influenced by non linear phenomena and depends on the organized structure of the system. The biological systems also have ordered metastable structures whose dimension are of the order of tens of nanometer and this could influence the behaviour of the electronic orbitals relative to the single molecules. The idea is that in a biological cell there is a large variety of low dimensional macromolecules (as, for instance, alpha-helical polypeptide proteins, actin filaments, etc.) whose structure is represented by the arrays of parallel quasi-one-dimensional (quasi-1D) polypeptide chains formed by periodically placed peptide groups along the chain of hydrogen bonds. The chains are characterized by the strong exchange and/or resonance interaction between the neighboring molecules, while the interaction between the chains is much weaker. From the point of view of electronic structure, these macromolecules are semiconductor like quasi-1D systems with the filled valence band and empty conduction band, separated by the gap of finite width. These characteristic properties favour the existence of coherent collective electron and exciton states, in general, and solitons in particular.
Obviously biosystems posses specific delayed photoluminescence whose properties differ significantly from those of similar devitalised organisms, and both are different from non-living systems and depend on the physical, chemical, physiological and other conditions and pre-history of the samples. All our experimental results show that the emission mechanism is influenced by non linear phenomena and depends on the organized structure of the system. The biological systems also have ordered metastable structures whose dimension are of the order of tens of nanometer and this could influence the behaviour of the electronic orbitals relative to the single molecules. The idea is that in a biological cell there is a large variety of low dimensional macromolecules (as, for instance, alpha-helical polypeptide proteins, actin filaments, etc.) whose structure is represented by the arrays of parallel quasi-one-dimensional (quasi-1D) polypeptide chains formed by periodically placed peptide groups along the chain of hydrogen bonds. The chains are characterized by the strong exchange and/or resonance interaction between the neighboring molecules, while the interaction between the chains is much weaker. From the point of view of electronic structure, these macromolecules are semiconductor like quasi-1D systems with the filled valence band and empty conduction band, separated by the gap of finite width. These characteristic properties favour the existence of coherent collective electron and exciton states, in general, and solitons in particular.
The concept of molecular solitons or 1D polaron-type states, which participate in charge and energy transport during the metabolic processes, was first suggested by A. Davydov and N. Kislukha. When in the electron transfer chain an extra electron is transferred from a donor molecule to one of the sub-units of the macromolecule, it affects the chain structure due to the exchange interaction with the nearest neighbours and electron-phonon interaction with hydrogen bonds along a chain. This results in the creation of the local distortion of the chain, which, in its turn, plays the role of the potential well for the electron itself and leads to the self-trapping (autolocalisation) of an electron. The suggested model assumes that the DL is connected with the formation and dissociation of non-linear coherent self-trapped (localised) electron states (solitons and electrosolitons) which are much more stable, in view of long duration of DL, than the corresponding delocalised states, and can be created, in low-dimensional macromolecular structures in the cell during the charge and energy transfer processes, by the pre-illumination of the sample. This model connecting this phenomenon with coherent eletron states in macromolecules in able to explain the long duration of DL and its main properties in biosystems, such as DL kinetcs satisfing an hyperbolic time dependence in a wide time inteval (I(t)=(a+bt)-α); same time trend for the various components of the emission spectrum; initial intensity od DL (I(t=0)) depending on the intensity of stimulating light in a nonlinear way.